Στο παρόν άρθρο θα γίνει μία σύντομη ανασκόπηση ορισμένων σημαντικών ανακαλύψεων και εξελίξεων όσον αφορά στις ασθένειες της μέλισσας (δημοσιευμένα 2019 και 2020). Όλα τα ευρήματα έχουν πρακτική σημασία για τον μελισσοκόμο, οπότε τα σχολιάζουμε.
Α΄ Αμερικάνικη Σηψηγονία
Ταυτοποιήθηκε το πέμπτο σε σειρά στέλεχος του βάκιλου της αμερικάνικης σηψηγονίας το οποίο ταξινομήθηκε μαζί με τα υπόλοιπα ήδη γνωστά στελέχη [1]. Το στέλεχος βρέθηκε σε ισπανικό μέλι και έλαβε την ονομασία ERIC-V. Παράλληλα, χαρτογραφήθηκε το DNA του μικροβίου για όλα τα γνωστά στελέχη. Τα στελέχη εξετάστηκαν εργαστηριακά ως προς τη ταχύτητα θανάτωσης της λάρβας (LD100), σε λάρβες οι οποίες μολύνθηκαν με 500 σπόρια η κάθε μία.
- Το πολύ συχνά απαντώμενο στέλεχος ERIC I έχει LD100 ~ 12 ημέρες
- Το συχνά απαντώμενο στέλεχος ERIC II, έχει LD100 ~ 7 ημέρες
- Τα σπάνια στελέχη ERIC III και IV έχουν LD100 ~3 ημέρες
- και τέλος το πολύ σπάνιο στέλεχος ERIC V το οποίο έχει LD100 ~ 3ημέρες
Ο τύπος ERIC I είναι ο βραδύτερα επιθετικός και ο συχνότερα απαντώμενος τύπος στα μελισσοκομεία δίνοντας τα κλασσικά συμπτώματα [2]. Ο τύπος ERIC II είναι μεσαίας φονικότητας και είναι σπανιότερος από τον πρώτο και δίνει πολλές φορές ψευδώς αρνητικά τέστ λόγω του χαμηλού αριθμού προσβεβλημένων κελιών τα οποία εμφανίζονται στα άρρωστα μελίσσια [2]. Η γενεαλογική ταξινόμηση του νεοανακαλυφθέντος και ERIC V (γοργά θανατηφόρου) δείχνει ότι αποτελεί κάποιο πρόγονο του τύπου ERIC II. Τα στελέχη ΙΙΙ, ΙV και V έχουν εκλείψει επιδημιολογικά. Είναι, επίσης, γνωστό ότι τα στελέχη διαφοροποιούνται ως προς τα συμπτώματα αλλά και ως προς το μεταβολικό προφίλ. Ακολουθεί η σχηματική απεικόνιση της παθογένειας της Αμερικάνικης Σηψηγονίας:
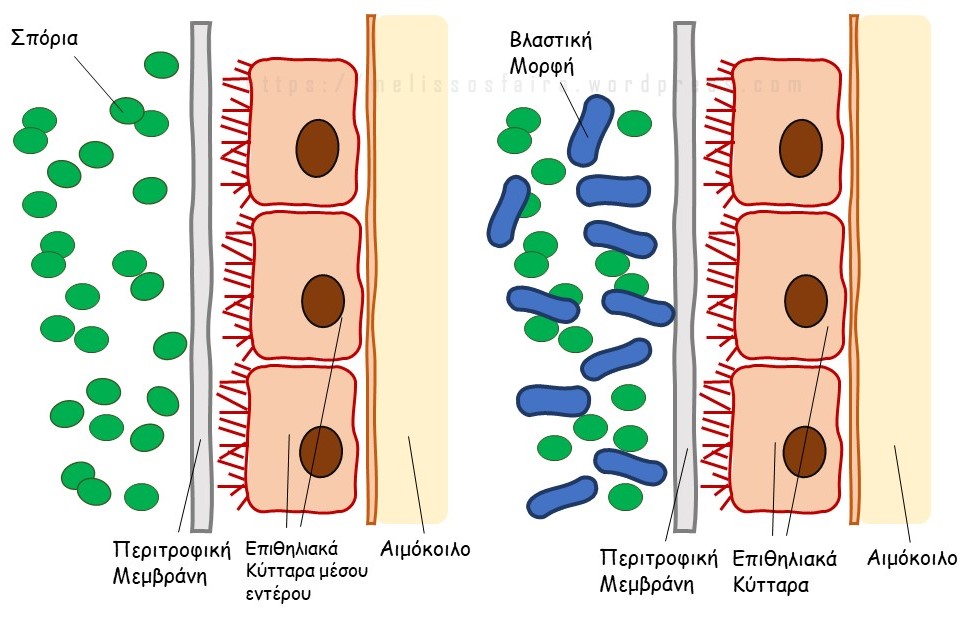
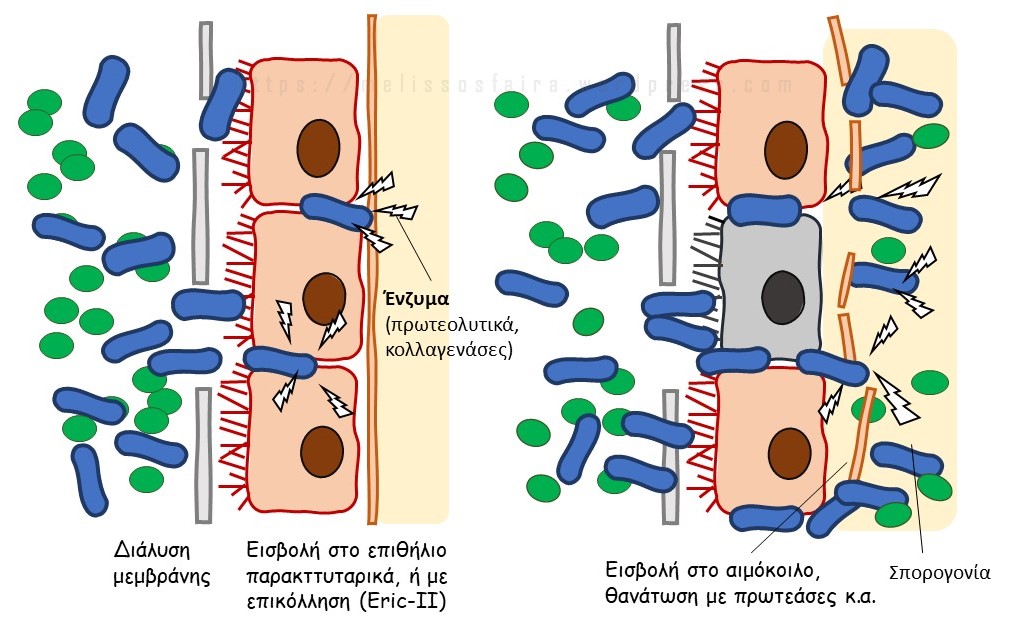
Η φάση της σήψης ξεκινάει με την εισβολή στο αιμόκοιλο και είναι η φάση κατά την οποία παράγονται τα περισσότερα σπόρια της Α.Σ. Να σημειωθεί ότι πολλά μέλια έχουν εκ του φυσικού τους σπόρια του βακτηρίου. Το βακτήριο είναι εξειδικευμένο στο να προσβάλλει μόνο τον γόνο της μέλισσας και το μέλι είναι ασφαλές προς ανθρώπινη κατανάλωση.
Σχολιασμός: Οι τύποι ERIC III, IV και V δεν παρατηρούνται στα μελίσσια, πιθανώς λόγω της πολύ γρήγορης θανάτωσης της λάρβας η οποία προκαλεί την άμεση απομάκρυνσή της από τις εργάτριες. Αυτά τα στελέχη του βακτηρίου δεν προλαβαίνουν να κάνουν μεγάλη σπορογονία γιατί το μελίσσι προλαβαίνει να κάνει εξυγίανση προτού σφραγιστεί ο γόνος.
Η επικράτηση του στελέχους ERIC I σε σχέση με το ERIC II δείχνει την εξελικτική τάση που έχει ακολουθήσει η νόσος. Τα βραδέως θανατηφόρα στελέχη τείνουν να ευνοούνται έναντι των γρήγορων και θανατηφόρων διότι είναι, μάλλον, προς όφελος του βακτηρίου να μην γίνεται εύκολα αντιληπτός ο προσβεβλημένος γόνος από τις εργάτριες μέλισσες. H ανακάλυψη καθώς και το προφίλ του νέου πέμπτου στελέχους (ERIC V) ενισχύει αυτήν την υπόθεση [1]. Η νόσος δείχνει να έχει προσαρμοστεί, κυρίως, στην εξελικτική πίεση που έχει ασκήσει η φυσική εξυγιαντική συμπεριφορά των Μελισσιών. Η χρήση αντιβιοτικών σε παγκόσμιο επίπεδο, πιθανόν να έχει επιτείνει το φαινόμενο. Όχι μόνο λόγω της γνωστής ανθεκτικότητας που έχει αναπτύξει το μικρόβιο στα αντιβιοτικά αλλά και εξαιτίας του παθογενικού προφίλ. Δηλαδή μιας «απαρατήρητης» εξέλιξης κατά τα πρώτα στάδια.
Για καλή μας τύχη τα αντιβιοτικά έχουν απαγορευτεί στην Ε.Ε., οπότε η χρήση τους έχει περιοριστεί δραστικά. Σε ορισμένες μελισσοκομικές επιχειρήσεις εμμένει ακόμα η απαρχαιωμένη πρακτική των «προληπτικών αντιβιώσεων» που αντιβαίνει σε κάθε γνωστή αρχή της βιολογίας, με τη μέλισσα να αντιμετωπίζεται σαν «μόνιμος ασθενής». Ουσιαστικά, με τις αντιβιώσεις επιχειρούν να κουκουλώσουν τα πολύ μεγάλα φορτία των σπορίων του βακτηρίου και να μετακυλήσουν το βαρύ κόστος της καταστροφής και αντικατάστασης του υλικού. Το αποτέλεσμα είναι όχι μόνο η κάλυψη αλλά ταυτόχρονα και η εύνοια της εξέλιξης της νόσου. Μιλάμε για αχρήστευση της γαστρεντερικής χλωρίδας των μελισσών από την δράση των αντιβιοτικών, κάτι που φαίνεται ότι δεν είναι σε θέση να προστατέψει ούτε τις επόμενες γενιές από τις λοιμώξεις. Τα αντιβιοτικά που χρησιμοποιούν σε ορισμένες περιπτώσεις στην Ασία είναι τα Νιτροφουράνια, παλαιότερα η Χλωροαμφενικόλη κ.α. Στη δύση είναι η Τερραμυκίνη (οξυτετρακυκλίνη), η Τυλοσίνη και παλαιότερα οι Σουλφοναμίδες [3]. Είναι δύσκολο να παρακολουθήσει κανείς τις τάσεις στη χρήση αντιβιοτικών μιας και τα αλλάζουν συχνά. Προ μερικών δεκαετιών, η επιστημονική ομοφωνία δεν καταδίκαζε τα αντιβιοτικά στη μελισσοκομία διότι θεωρούσαν ότι δρούσαν επιλεκτικά μόνο στις βλαστικές μορφές και έτσι δεν ανθεκτικοποιούσαν τα βακτήρια. Η εξέλιξη των πραγμάτων έχει διαψεύσει αυτή τη θεώρηση γιατί πλέον όλα τα παθογόνα στη μελισσοκομία έχουν αποκτήσει κάποιας μορφής ανθεκτικότητα.
Β΄ Νοσεμίαση
Έγινε ανασκόπηση της βιβλιογραφίας για τη σχέση Νοσεμίασης και Δυσεντερίας στις μέλισσες από τον Randy Oliver μέσα από μία σειρά άρθρων του [5]. Δεν βρέθηκε να έχει καταγραφεί κάποια αιτιακή σχέση μεταξύ τους. Δηλαδή, η Νοσεμίαση Apis και Ceranae δεν προκαλούν δυσεντερία. Η ενοχοποίηση της Νοσεμίασης για το συμπτώμα της διάρροιας αναπαράγεται για χρόνια σε εργασίες και σε μελισσοκομικές συζητήσεις, χωρίς μέχρι σήμερα να έχει επιβεβαιωθεί πειραματικά. H συσχέτιση πιθανώς να ξεκίνησε από μία εργασία του L. Bailey το 1967 και έκτοτε αναπαράχθηκε χιλιάδες φορές (αν και ούτε ο Bailey υποστήριξε την αιτιακή σχέση μεταξύ των δύο) [6].
Τα τελευταία χρόνια, η υπόθεση αυτή έχει εξεταστεί και έχει αμφισβητηθεί πολλές φορές, χωρίς όμως να προκαλέσει αλλαγές και διορθώσεις μεγάλης κλίμακας. Για παράδειγμα, ούτε στην παλαιότερη ελληνική βιβλιογραφία αναγνωρίζεται η Νοσεμίαση Apis ως υπεύθυνη για τη δυσεντερία, παρά μόνο στο ότι μπορεί να την επιτείνει όταν ήδη υπάρχει (Υφαντίδης 1995, 2011) [14].
Ο Randy Oliver έδωσε μια ενδιαφέρουσα εκδοχή για την αιτία της χειμερινής διάρροιας στις μέλισσες σε μία σειρά άρθρων [7], [8]. Σύμφωνα με αυτή, η δυσεντερία είναι σύμπτωμα της κακής ρύθμισης του νερού που παράγεται κατά τη παρατεταμένη διαχείμαση του σμήνους στην κατάσταση της μελισσόσφαιρας.
Σχολιασμός. Η Νοσεμίαση οφείλεται σε μικροσπορίδιο (είδος μύκητα) το οποίος προσβάλλει το μεσέντερο της μέλισσας (το ανθρώπινο ανάλογο του στομαχιού και όχι του εντέρου). O στόχος του μύκητα είναι τα επιθηλιακά κύτταρα του στομαχιού. Τα κύτταρα αυτά, στα προχωρημένα στάδια της νόσου, καταστρέφονται εντελώς (διαρρηγνύονται) και επομένως η μέλισσα δεν μπορεί πλέον να απορροφήσει θρεπτικά συστατικά. Όμως, η σπορογονία γίνεται στο στομάχι ασχέτως αν ο πεπτικός σωλήνας καταλήγει να είναι γεμάτος σπόρια. Πέραν του στομαχιού, δεν είναι γνωστές κάποιες ενέργειες των σπορίων στο απευθυσμένο οι οποίες θα προκαλούσαν διάρροια πχ. έκκριση τοξινών κ.α. Φυσικά η δυσεντερία γίνεται να διασπείρει σπόρια εντός της κυψέλης και να αυξήσει το φορτίο. Αξίζει να σημειωθεί ότι τα δύο παθογόνα N. Apis και N. Ceranae διαφέρουν πολύ, τόσο σε συμπτωματολογία όσο και γενεαλογικά. H Νοσεμίαση Ceranae μπορεί και προσβάλλει με μικρό βαθμό και άλλα όργανα όπως οι υποφαρυγγικοί αδένες, το λιπόσωμα κ.α.[9], [10]. Η Nosema Ceranae έχει μεγαλύτερη σχέση με την Νοσεμίαση της Σφήκας παρά με την Nosema Apis [10]
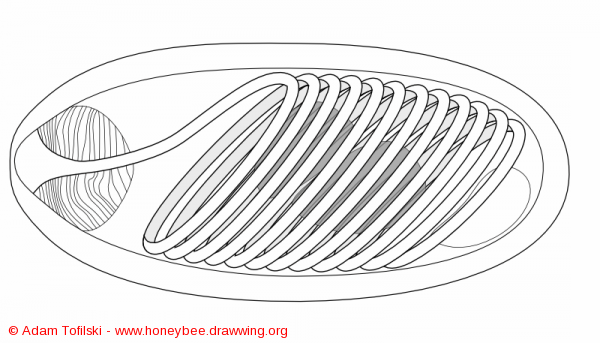
Μη βλαστική μορφή του Mικροσπορίδιου[11]. Όταν βλαστήσει, λαμβάνει σωληνωτή μορφή με την οποία τρυπάει το επιθηλιακό κύταρρο του Μεσέντερου (Στομάχι). Κατόπιν, ρίχνει μέσα το σπορόπλασμα και ξεκινάει ο πολλαπλασιασμός. Μία παρόμοια παρανόηση υπάρχει και όταν παρατηρούνται μέλισσες να σέρνονται μπροστά στις εισόδους της κυψέλης με τα φτερά ασυντόνιστα αδυνατώντας να πετάξουν, κάτι που ερμηνεύεται ως «σύμπτωμα Νοσεμίασης». Στην πραγματικότητα, τα συμπτώματα προκαλούνται από τον Ιό της Χρόνιας Παράλυσης (CBPV) που μεταφέρεται στη μέλισσα από τη βαρρόα. Αντιθέτως, η παθογένεια της Νοσεμίασης προκαλεί στις μέλισσες αύξηση της κατανάλωση τροφής (λόγω αδυναμίας απορρόφησης θρεπτικών) και τελικά τις οδηγεί στο να χάνονται μακρυά από την κυψέλη, όχι στις εισόδους. Η νοσεμίαση πολλές φορές συνυπάρχει στις μέλισσες που νοσούν από τον Ιό, χωρίς όμως να είναι η αιτία της παράλυσης. Μάλιστα, έχει παρατηρηθεί εργαστηριακά συνέργεια της Nosema Ceranae με τον Ιό της Χρόνιας Παράλυσης [12]. Επομένως, η κατάσταση είναι σύνθετη. Το σίγουρο λάθος είναι όταν παρατηρούνται παράλυτες μέλισσες στις εισόδους να λαμβάνονται μέτρα για τη Νοσεμίαση, αντί να ληφθούν μέτρα για την πρωτογενή αιτία, τη βαρρόα.
Γ΄ Ανθεκτικότητα στη Βαρρόα
Ανακαλύφθηκε ένα γενετικό χαρακτηριστικό το οποίο προσδίδει ανθεκτικότητα στη βαρρόα σε δύο πληθυσμούς, έναν στη Τουλούζη (Γαλλία) και έναν στο Νησί του Gotland (Σουηδία) [13]. Και οι δύο πληθυσμοί παρόλο που απέχουν μεγάλη απόσταση τυγχάνει να έχουν ένα παρόμοιο μηχανισμό ανθεκτικότητας. Πως λειτουργεί ο μηχανισμός; Η βαρρόα δεν έχει τη δυνατότητα να συνθέσει κάποιες ορμόνες κι έτσι είναι αναγκασμένη να τις παίρνει αυτούσιες από τη μέλισσα ξενιστή. Μια τέτοια απαραίτητη ορμόνη είναι η εκδυσόνη. Η εκδυσόνη είναι (μια στεροειδής προορμόνη) η οποία χρησιμοποιείται από το έντομο στις διαδικασίες της έκδυσης και της μεταμόρφωσης. Τρία συνολικά γονίδια ανθεκτικότητας βρέθηκαν σχετιζόμενα με την εκδυσόνη (δύο στον πληθυσμό του Gotland και ένα στον πληθυσμό της Τουλούζης). Οι διαταραχές στην πρόσληψη εκδυσόνης από τη βαρρόα-γενάρχη μπορεί να οδηγήσουν είτε στο να ματαιώσει η βαρρόα την ωοτοκία είτε να μην μπορούν να αναπτυχθούν νέα βαρρόα, μιας και η εκδυσόνη δίνει το έναυσμα για την σύνθεση της βιτελλογενίνης. Από το κελί τελικά βγαίνει ζωντανή μόνο η μάνα βαρρόα κάτι που οδηγεί προοδευτικά στον περιορισμό του πληθυσμού της βαρρόα.
Σχολιασμός: Σε προηγούμενο άρθρο αναλύθηκε το θέμα της ανθεκτικότητας. Κάποιοι πρόγονοι των συγκεκριμένων πληθυσμών είχαν την τύχη να διαθέτουν τα λιγοστά γονίδια που μπλοκάρουν την αναπαραγωγή του βαρρόα. Ένα χαρακτηριστικό το οποίο οφείλεται σε ένα- δύο αλληλόμορφα γονίδια σημαίνει ότι κληρονομείται εύκολα, επομένως η εξάπλωσή του υπό την πίεση του επιθετικού παρασιτισμού ήταν θέμα χρόνου.

Η έκφραση των εν λόγω γονιδίων γίνεται περισσότερο στον Κηφήνα και έτσι ο πληθυσμός της βαρρόα μένει στάσιμος, ή φθίνει για το διάστημα που το μελίσσι εκτρέφει κηφήνες. Τα γονίδια εκφράζονται περισσότερο στον κηφήνα παρά στην εργάτρια, καθώς δύναται να μην επικρατήσουν λόγω ετεροζυγωτίας.Η έκπληξη της ανακάλυψης ήταν ότι ενώ οι έρευνες κατευθύνονται προς τη συμπεριφορική ανθεκτικότητα των μελισσών με εξεζητημένες ιδιότητες αυτοκαθαρισμού (VSH), τελικά προέκυψε μια ξερή βιολογική λειτουργία την οποία ελάχιστοι μπορούσαν να φανταστούν. Δεν γίνεται να μη σχολιαστεί η καιροσκοπική φιλολογία που ξεφύτρωσε όταν παρουσιάστηκαν οι ανθεκτικοί πληθυσμοί στη βαρρόα (Gotland, Τουλούζη κτλ). Χωρίς κανείς να είναι σε θέση να γνωρίζει τι ακριβώς έχει συμβεί, αναπτύχθηκαν πολλές αυθαίρετες θεωρίες για το πως θα φτιαχτούν τα ανθεκτικά μελίσσια στη βαρρόα. Έγινε θεωρητικοποίηση των γεγονότων («κοιτάχτε πως οι μέλισσες έδωσαν μόνες τους τη λύση»), θεωρητικοποίηση των μεθόδων («για να επιταχύνουμε τη φυσική επιλογή ακολουθούμε τον Χ τρόπο»). Επιπλέον, ολη αυτή η φιλολογία όπλισε επικίνδυνα δόγματα αφάρμακης μελισσοκομίας.
Συμπερασματικά, πρόκειται για αισιόδοξη ανακάλυψη μιας και ανοίγονται μια σειρά από πλεονεκτήματα:
- Το χαρακτηριστικό κληρονομείται εύκολα, εν αντιθέσει με τα γονίδια της συμπεριφορικής ανθεκτικότητας (VHS).
- Εκφράζεται και στον κηφήνα, εκτός από την εργάτρια.
- Διατηρείται σε βάθος γενεών, χωρίς να επηρρεάζεται ιδιαίτερα από την πολυανδρία της μέλισσας.
- Ο εντοπισμός ανθεκτικών μελισσιών μπορεί να γίνει με εργαστηριακό τεστ αντί για περίπλοκες και επίπονες διαδικασίες στον αγρό.
Δεν πρέπει να ξεχνιέται, όμως, ότι η εν λόγω ανθεκτικότητα βασίζεται ουσιαστικά σε ελλατωματική λειτουργία των μηχανισμών παραγωγής εκδυσόνης με πιθανό αντίκτυπο στην ολοκλήρωση της ανάπτυξης του γόνου. Μιλάμε για ελαφρώς ελλατωματική μέλισσα η οποία όμως δεν καταβάλλεται από το παράσιτο.
Γιώργος Μήτσικας (Χημικός, Ερασιτέχνης Μελισσοκόμος)
Μάρτιος 2021
ΒΙΒΛΙΟΓΡΑΦΙΑ- ΠΑΡΑΠΟΜΠΕΣ
[1] Beims, H., Bunk, B., Erler, S., Mohr, K. I., Spröer, C., Pradella, S., … & Steinert, M. (2020). Discovery of Paenibacillus larvae ERIC V: Phenotypic and genomic comparison to genotypes ERIC I-IV reveal different inventories of virulence factors which correlate with epidemiological prevalences of American Foulbrood. International Journal of Medical Microbiology, 310(2), 151394.
[2] Infection of honey bees with Paenibacillus larvae (American Foolbrood). OIE Terrestrial Manual 2018, Chapter 9.2.
[3] Djukic, M., Brzuszkiewicz, E., Fünfhaus, A., Voss, J., Gollnow, K., Poppinga, L., … & Daniel, R. (2014). How to kill the honey bee larva: genomic potential and virulence mechanisms of Paenibacillus larvae. PloS one, 9(3), e90914.
[4] Reybroeck, W., Daeseleire, E., De Brabander, H. F., & Herman, L. (2012). Antimicrobials in beekeeping. Veterinary microbiology, 158(1-2), 1-11.
[5] Randy Oliver (2019). The Nosema Problem: Part 1 It’s “Common Knowledge” That Nosema Causes Dysentery But Is It Actually True? American Bee Journal May 2019.
[6] Bailey L (1967): Nosema apis and dysentery of the honey bee. J Apicultural Res 6: 121-125.
[7] Randy Oliver (2019). The Nosema Problem: Part 7a. The Causes Of Dysentery In Honey Bees Part 1. American Bee Journal December 2019
[8] Randy Oliver (2020). The Nosema Problem: Part 7b. The Causes Of Dysentery In Honey Bees: Part 2. American Bee Journal January 2020
[9] Goblirsch, M. (2018). Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie, 49(1), 131-150.
[10] Chen YP, Evans JD, Murphy C, Gutell R, Zuker M, et al. (2009) Morphological, molecular, and phylogenetic characterization of Nosema ceranae, a microsporidian parasite isolated from the European honey bee, Apis mellifera. J Euk Microbiol 56: 142–147.
[11] http://honeybee.drawwing.org/book/nosema (For Non Commercial use).
[12] Toplak, Ivan & Jamnikar ciglenecki, Urska & Aronstein, Katherine & Gregorc, Aleš. (2013). Chronic Bee Paralysis Virus and Nosema ceranae Experimental Co-Infection of Winter Honey Bee Workers (Apis mellifera L.). Viruses. 5. 2282-97. 10.3390/v5092282.
[13] A gene for resistance to the Varroa mite (Acari) in honey bee (Apis mellifera) pupae
Mol. Ecol., 28 (2019), pp. 2958-2966
[14] ΠΑΘΗΣΕΙΣ ΤΟΥ ΜΕΛΙΣΣΙΟΥ- Μη συμβατικές μέθοδοι αντιμετώπισης τους – Μιχαήλ Δ. Υφαντίδης. Μάιος 2011. Εκδ. Μελισσοκομική Επιθεώρηση

Σχολιάστε